- Denaturation at 94°C :
- Annealing at 50°C :
- extension at 60°C :
During the denaturation, the double strand melts open to single stranded DNA, all enzymatic reactions stop (for example : the extension from a previous cycle).
In sequencing reactions, only one primer is used, so there is only one strand copied (in PCR : two primers are used, so two strands are copied). The primer is jiggling around, caused by the Brownian motion. Ionic bonds are constantly formed and broken between the single stranded primer and the single stranded template. The more stable bonds last a little bit longer (primers that fit exactly) and on that little piece of double stranded DNA (template and primer), the polymerase can attach and starts copying the template. Once there are a few bases built in, the ionic bond is so strong between the template and the primer, that it does not break anymore.
This is the ideal working temperature for the polymerase (normally it is 72 °C, but because it has to incorporate ddNTP's which are chemically modified with a fluorescent label, the temperature is lowered so it has time to incorporate the 'strange' molecules. The primers, where there are a few bases built in, already have a stronger ionic attraction to the template than the forces breaking these attractions. Primers that are on positions with no exact match, come loose again and don't give an extension of the fragment.
The bases (complementary to the template) are coupled to the primer on the 3'side (adding dNTP's or ddNTP's from 5' to 3', reading from the template from 3' to 5' side, bases are added complementary to the template)
When a ddNTP is incorporated, the extension reaction stops because a ddNTP contains a H-atom on the 3rd carbon atom (dNTP's contain a OH-atom on that position). Since the ddNTP's are fluorescently labeled, it is possible to detect the color of the last base of this fragment on an automated sequencer.
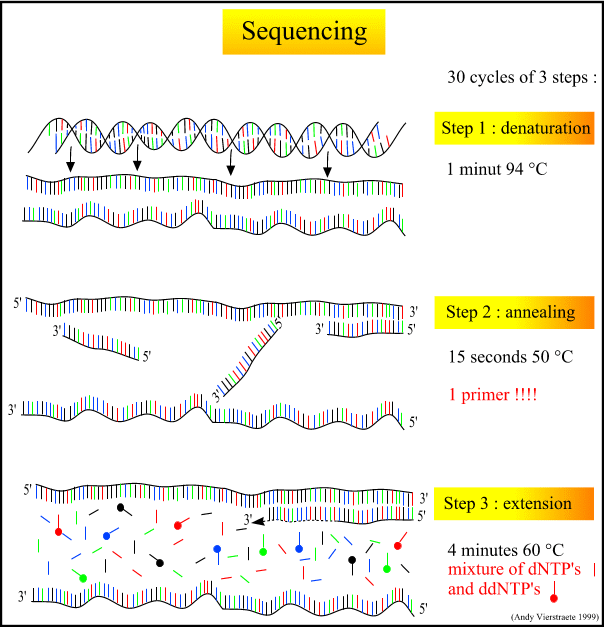
Figure 7 : The different steps in sequencing. (pdf file of this picture) Animated picture of sequencing (344 kB) Because only one primer is used, only one strand is copied during sequencing, there is a linear increase of the number of copies of one strand of the gene. Therefore, there has to be a large amount of copies of the gene in the starting mixture for sequencing. Suppose there are 1000 copies of the wanted gene before the cycling starts, after one cycle, there will be 2000 copies : the 1000 original templates and 1000 complementary strands with each one fluorescent label on the last base, after two cycles, there will be 2000 complementary strands, three cycles will result in 3000 complementary strands and so on.
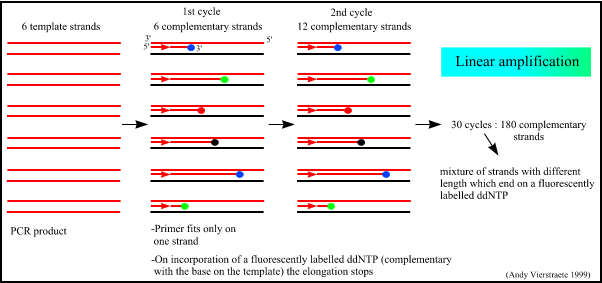
Figure 8 : The linear amplification of the gene in sequencing.
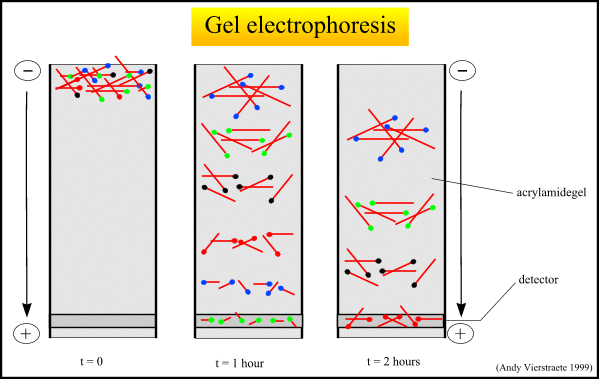
Figure 9 : The separation of the molecules with electrophoresis.(pdf file of this picture) Animated picture of gel electrophoresis (159 kB)
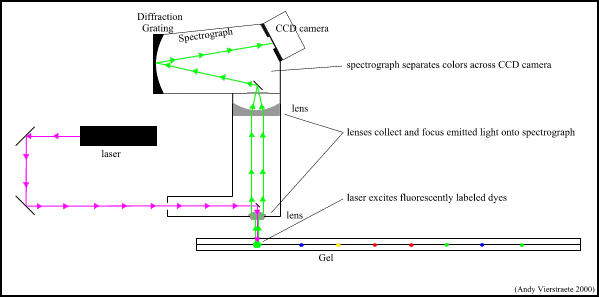
Figure 10 : The scanning and detection system on the ABI Prism 377 sequencer. (pdf file of this picture) Animated picture of scanning and detection system (182 kB)
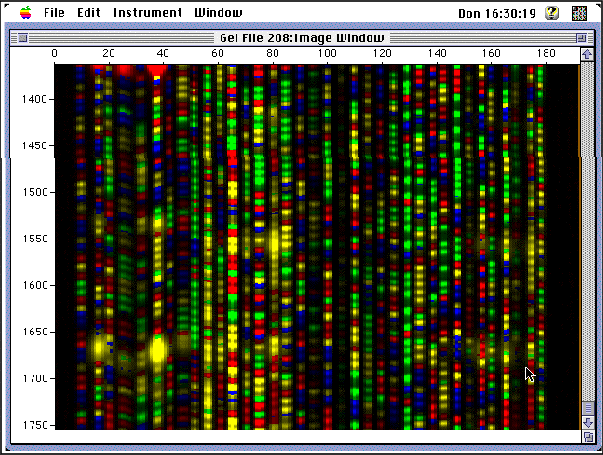
Figure 11 : A snapshot of the detection of the molecules on the sequencer.
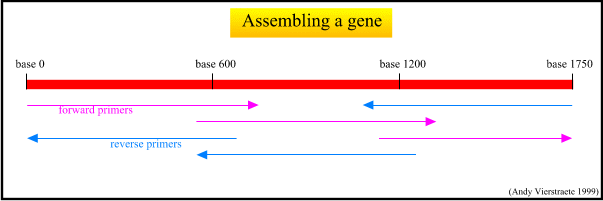
Figure 12 : The assemblage of the gene.